
通过可解释的人工智能平台 AgeXtend 发现老年保护剂
作者:Ahuja, Gaurav
数据可用性
支持研究结果的所有数据均可根据要求从通讯作者处获得。有关训练数据集的信息可通过 Zenodo 访问:https://doi.org/10.5281/zenodo.8378959(参考。54)。补充表、有关训练数据集的附加数据以及酵母年代测定的内源代谢物可通过 Zenodo 获取,网址为:https://doi.org/10.5281/zenodo.8379479(参考。55)。该研究利用了 20 个数据库的数据,包括: AfroDb51、老化图集 (v.1.0) (https://ngdc.cncb.ac.cn/aging), ChEMBL31 (https://www.ebi.ac.uk/chembl), 化学桥 (https://chembridge.com), ChemDiv BBlocks (https://www.chemdiv.com), CMNPD (v.1.0) (https://www.cmnpd.org)、DSSTox (v.2021) (https://comptox.epa.gov/dashboard),杜克博士的植物化学和民族植物学数据库(https://phytochem.nal.usda.gov),大肠杆菌代谢组 (v.2.0) (https://ecmdb.ca), FooDB (v.1.0, 预发布) (www.foodb.ca), GDB13 (https://gdb.unibe.ch), gutMGene (人类)52、HMDB (v.5.0) (https://hmdb.ca), IMPPAT 2.0 (v.2.0) (https://cb.imsc.res.in/imppat), 代谢时代 (https://www.metaboage.info)、MolData (v.1.1.0) (https://github.com/LumosBio/MolData), PubChem(2022 年 10 月 21 日下载)(https://pubchem.ncbi.nlm.nih.gov),药物再利用中心 (RepoHub)(2020 年 3 月 24 日发布)(https://repo-hub.broadinstitute.org/repurifying)、YMDB (v.2.0) (https://www.ymdb.ca) 和 ZINC12 (https://zinc12.docking.org)。
代码可用性
AgeXtend 的 Python 包通过 pip 提供:https://pypi.org/project/AgeXtend。AgeXtend 对学术机构免费;但是,商业用途必须有商业许可证密钥。AgeXtend 的源代码可在项目 GitHub 上找到:https://github.com/the-ahuja-lab/AgeXtend也可以通过 Zenodo 在https://doi.org/10.5281/zenodo.8378959(参考。54)。AgeXtend 工作流程也可通过 CodeOcean 获得,网址为https://codeocean.com/capsule/0693261/tree。参考
López-Otín, C.、Blasco, M. A.、Partridge, L.、Serrano, M. 和 Kroemer, G. 衰老的标志。
细胞153 ,1194-1217 (2013)。文章
一个 考研一个 考研中心一个 谷歌学术一个 López-Otín, C.、Blasco, M. A.、Partridge, L.、Serrano, M. 和 Kroemer, G. 衰老的标志:膨胀的宇宙。细胞
186,243–278 (2023)。 文章一个
考研一个 谷歌学术一个 Solovev, I., Shaposhnikov, M. & Moskalev, A. 人类生物年龄估计的多组学方法。机甲。
老化开发。185 ,111192(2020)。文章
一个 中科院一个 考研一个 谷歌学术一个 DönertaÅ,H.M.,Fuentealba,M.,Partridge,L. 和 Thornton,J.M. 在计算机中识别潜在的衰老调节药物。趋势内分泌。
代谢物。30 ,118–131 (2019)。文章
老化研究。牧师。 64,101154(2020)。
Prieto-Oliveira,P.端粒酶激活在衰老或退行性疾病治疗中的作用:系统评价。摩尔。细胞。生物化学。 第476章,599–607 (2021)。
莫雷尔,J.-D。等人。小鼠衰老和新陈代谢的金属景观。纳特。交流。 13,607(2022)。
Barzilai, N.、Huffman, D. M.、Muzumdar, R. H. 和 Bartke, A. 代谢途径在衰老中的关键作用。糖尿病 61,1315 - 1322 (2012)。
Hernández-Camacho, J. D.、Bernier, M.、López-Lluch, G. 和 Navas, P. 辅酶 Q10 补充剂对衰老和疾病的影响。正面。生理学。 9,44(2018)。
Hipkiss,A. R. 衰老、蛋白毒性、线粒体、糖化、NAD 和肌肽:可能的相互关系和氧悖论的解决。正面。衰老神经科学。 2,10(2010)。
辛格,P.等人。牛磺酸缺乏是衰老的驱动因素。科学 380, eabn9257 (2023)。
Hosseini, L.、Vafaee, M. S.、Mahmoudi, J. 和 Badalzadeh, R. 烟酰胺腺嘌呤二核苷酸成为衰老和缺血性疾病的治疗靶点。生物老年学 20,381–395 (2019)。
Correia-Melo,C. 等人。细胞与细胞之间的代谢物交换创造了一个有利于生存的代谢环境,可以延长寿命。细胞186 ,63–79.e21 (2023)。文章
一个 中科院一个 考研一个 谷歌学术一个 DönertaÅ,H. M.,Fuentealba Valenzuela,M.,Partridge,L. 和 Thornton,J. M. 基于基因表达的药物重新利用以靶向衰老。老化细胞
17 号,e12819(2018)。 文章一个
考研一个 考研中心一个 谷歌学术一个 Mittal, A. 和 Ahuja, G. 推进化学致癌性预测模型:机遇与挑战。趋势药理学。
科学。44 ,400–410 (2023)。文章
一个 中科院一个 考研一个 谷歌学术一个 Fernández-Torras, A.、Comajuncosa-Creus, A.、Duran-Frigola, M. 和 Aloy, P。通过分子描述符连接化学和生物学。电流。
意见。化学。生物。 66,102090(2022)。
巴拉多,D.等人。与衰老相关的药物的 DrugAge 数据库。老化细胞 16,594–597 (2017)。
Moskalev, A.、Chernyagina, E.、Kudryavtseva, A. 和 Shaposhnikov, M. Geroprotectors:统一的概念和筛选方法。老化问题。 8,354–363 (2017)。
Ye,X.,Linton,J.M.,Schork,N.J.,Buck,L.B. 和 Petrascheck,M. 延长寿命的药理学网络秀丽隐杆线虫。老化细胞 13,206–215 (2014)。
Kornfeld, K. 和 Evason, K. 抗惊厥药物对寿命的影响。拱。尼罗尔. 63,491–496 (2006)。
亨特,P.R.等人。延长寿命线虫通过通过应激兴奋机制起作用的萘醌。公共图书馆一号 6,e21922 (2011)。
Evason, K.、Huang, C.、Yamben, I.、Covey, D. F. 和 Kornfeld, K. 抗惊厥药物可延长蠕虫寿命。科学 307,258–262 (2005)。
Moriwaki, H.、Tian, Y.-S.、Kawashita, N. 和 Takagi, T. Mordred:分子描述符计算器。J.Cheminform。 10,4(2018)。
阿克森,S.D.等人。三维分子结构的简单表示。J. Med。化学。 60,7393–7409 (2017)。
拉姆桑达尔,B. 等人。DeepChem:使药物发现、量子化学的深度学习民主化。GitHub github.com/deepchem/deepchem(2019)。
贝尔托尼,M.等人。未表征的化合物的生物活性描述符。纳特。交流。 12,3932(2021)。
Hubbard, B. P. & Sinclair, D. A. 用于治疗衰老和年龄相关疾病的小分子 SIRT1 激活剂。趋势药理学。科学。 35,146–154 (2014)。
Kim, M. E.、Kim, D. H. 和 Lee, J. S. 转录因子作为与年龄相关的疾病和癌症中天然化合物的靶标:潜在的治疗应用。国际。J.莫尔。科学。 23,13882(2022)。
Fuhrmann-Stroissnigg, H., Niedernhofer, L. J. & Robbins, P. D. Hsp90 抑制剂作为延缓健康衰老的药物。细胞周期 17 号,1048–1055 (2018)。
Nieto-Torres, J. L. & Hansen, M. 巨自噬和衰老:细胞回收对健康和长寿的影响。摩尔。医学方面。 82,101020(2021)。
Kursa, M. B. 和 Rudnicki, W. R. 使用 boruta 包进行功能选择。J. 统计。软件。 36,1-13 (2010)。
Wassermann,A.M. 等人。暗化学物质是药物先导化合物发现的一个有希望的起点。纳特。化学。生物。 11,958–966 (2015)。
Mohammed, I.、Hollenberg, M. D.、Ding, H. 和 Triggle, C. R. 对二甲双胍是一种假定的抗衰老药物可增强健康寿命和延长寿命的证据进行严格审查。正面。内分泌。 12,718942(2021)。
Reymond, J.-L.、Ruddigkeit, L.、Blum, L. 和 van Deursen, R. 化学空间的枚举。电线计算。摩尔。科学。 2,717–733 (2012)。
Panyard, D. J.、Yu, B. 和 Snyder, M. P. 人类衰老的代谢组学:进展、挑战和机遇。科学。副词。 8,eadd6155 (2022)。
Ghosh, T. S.、Shanahan, F. 和 OâToole, P. W. 改进健康微生物组的定义,促进健康老龄化。纳特。老化 2,1054 - 1069 (2022)。
陈,Y.等人。将长寿人群的微生物群移植到小鼠体内可降低与衰老相关的指数并转移有益细菌。老化 12,4778–4793 (2020)。
海因肯,A.等人。7,302 种人类微生物的基因组规模代谢重建,用于个性化医疗。纳特。生物技术。 41,1320 - 1331 (2023)。
宋,J.等人。人类肠道微生物群的全球代谢相互作用网络,用于特定环境的群落规模分析。纳特。交流。 8,15393(2017)。
Ghosh, T. S.、Das, M.、Jeffery, I. B. 和 OâToole, P. W. 根据年龄进行调整可以提高对多种疾病中肠道微生物组变化的识别。电子生活 9,e50240 (2020)。
张,L.等人。细胞衰老:衰老和疾病的关键治疗靶点。J.克林。投资。 132,e158450 (2022)。
Ozsvari, B.、Nuttall, J. R.、Sotgia, F. 和 Lisanti, M. P. 阿奇霉素和罗红霉素定义了一个针对衰老人类成纤维细胞的新“抗衰老”药物家族。老化 10,3294–3307 (2018)。
楠本,D.等人。基于深度学习的形态学衰老评分进行抗衰老药物筛选。纳特。交流。 12,257(2021)。
Oeppen, J. 和 Vaupel, J. W. 人口统计学。打破了预期寿命的限制。科学 296,1029-1031 (2002)。
Crimmins,E.M.《寿命和健康寿命:过去、现在和未来》。老年学家 55,901–911 (2015)。
Wong, F.、Omori, S.、Donghia, N. M.、Zheng, E. J. 和 Collins, J. J. 通过深度神经网络发现小分子 senolytics。纳特。老化 3,734–750 (2023)。
Bradshaw,P.C. 细胞质和线粒体 NADPH 耦合氧化还原系统在衰老调节中的作用。营养素 11,504(2019)。
Schyman, P.、Liu, R.、Desai, V. 和 Wallqvist, A. 用于 ADMET 预测的 vNN Web 服务器。正面。药理学。 8,889(2017)。
Blagus, R. & Lusa, L. SMOTE,用于高维类不平衡数据。BMC 生物信息。 14,106(2013)。
Liu, T.、Lin, Y.、Wen, X.、Jorrisen, R. N. 和 Gilson, M. K. BindingDB:实验确定的蛋白质-配体结合亲和力的可通过网络访问的数据库。核酸研究。 35,D198→D201 (2007)。
Ntie-Kang,F.等人。AfroDb:来自非洲药用植物的精选高效且多样化的天然产物库。公共图书馆一号 8,e78085 (2013)。
程,L.等人。gutMGene:肠道微生物靶基因和微生物代谢物的综合数据库。核酸研究。 50,D795 – D800 (2022)。
Aging Atlas Consortium Aging Atlas:衰老生物学的多组学数据库。核酸研究。 49、D825 — D830 (2021)。
阿罗拉,S.等人。通过可解释的基于人工智能的平台 AgeXtend(训练数据集/代码)发现老年保护剂。泽诺多 https://doi.org/10.5281/zenodo.8378959(2023)。
阿罗拉,S.等人。通过基于人工智能的可解释平台 AgeXtend(附加数据)发现老年保护剂。泽诺多 https://doi.org/10.5281/zenodo.8379479(2023)。
致谢
我们感谢 IIIT-Delhi 的 IT-HelpDesk 团队提供计算资源方面的协助。我们感谢 G.A. 实验室的所有成员在该项目的各个阶段做出的智力贡献。我们感谢 K. Sriram 在数学计算方面提供的帮助。我们还感谢 V. K. Yenamandra 分享人类成纤维细胞,感谢 J. Tayal 和 A. Mehta 提供细胞培养设施,感谢 A. Mukhopadhyay 分享线虫菌株。G.A. 实验室得到了印度政府科学技术部生物技术司、科学与工程研究委员会的启动研究补助金(编号 SRG/2020/000232)、青年研究员计划的支持欧洲分子生物学组织 (EMBO) 和 IHUB Anubhuti 的研究资助(项目资助/23)以及 Indraprastha 信息研究所的校内启动资助技术-德里。印度科学技术部的 INSPIRE 教员资助为 D. Sengupta 实验室提供资金。我们衷心感谢 TCS 基金会赞助 S.A. 作为其 TCS 研究学者计划的一部分。
道德声明
利益竞争
作者声明没有竞争利益。
同行评审
同行评审信息
自然老化感谢 Peter Fedichev 和其他匿名审稿人对这项工作的同行评审所做的贡献。
附加信息
出版商备注施普林格·自然对于已出版的地图和机构隶属关系中的管辖权主张保持中立。
扩展数据
扩展数据 图 1 示意图详细说明了构建 AgeXtend 的 Geroprediction 模块所涉及的步骤。
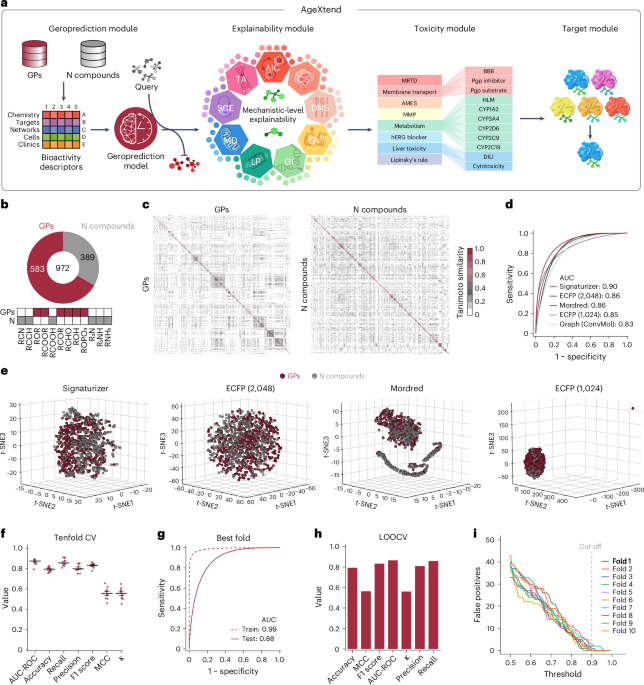
(一个) 冲积图显示了用于训练 AgeXtend 的毒性、Geroprediction 和可解释性模块中的各种模型的数据集的主要来源。(乙) 示意图描述了为 AgeXtend 构建 Geroprediction 模块最终模型所涉及的所有关键步骤。值得注意的是,四种不同类型的特征用于训练化合物的分子表示。输入数据集被随机分为训练数据集(75%)和测试数据集(25%),并使用 SMOTE 算法处理类别不平衡。选择最佳特征类型(生物活性)后,使用五重交叉验证策略进一步评估模型性能,并通过超参数调整进一步调整最佳分类器(SVM)以增强性能。使用 Boruta 进行特征选择。100% 的数据用于构建具有最佳性能超参数的最终模型。(c) 散点区间图(n=5 倍)描绘了在 5 倍交叉验证期间使用不同特征构建的指定模型的性能。须线代表 ± SE,粗体水平线表示平均值。模型性能使用接收器工作特性曲线下面积 (AUC-ROC) 和准确度来衡量。
扩展数据 图 2 Geroprotector 模型的预测性能。
(一个) 散点区间图(n=5 倍)描绘了使用接受者操作特征曲线下面积 (AUC-ROC) 曲线、准确度、F1 分数、精确度、召回率、Cohen 评估的指示模型性能。5 倍交叉验证期间的 s kappa 和 Matthew 相关系数 (MCC)。须线代表 ± SE,粗体水平线表示平均值。值得注意的是,模型是使用经过实验验证的老年保护剂和中性化合物作为输入的指定分类器建立的。测试了以下分类器:逻辑回归 (LR)、支持向量机 (SVM)、梯度提升分类器 (GBC)、随机森林 (RF)、高斯朴素贝叶斯 (GNB)、K 最近邻 (KNN)、额外树分类器 (ET)、随机梯度下降(SGD)、XGBoost(XGB)和多层感知器(MLP)。(乙) 散点区间图描绘了使用 geroprotector/neutral 数据集构建的 SVM 模型的留一交叉验证 (LOOCV) 的训练性能指标(n==972 倍)。须线代表 ± SE,粗体水平线表示平均值。(c) AUC(曲线下面积)图,描述留一交叉验证 (LOOCV) 的测试性能。(d) 示意图描述了构建 AgeXtend 的可解释性和毒性模块模型所涉及的所有关键步骤。值得注意的是,基于签名的生物活性描述符用于训练化合物的分子表示。输入数据集被随机分为训练数据集(75%)和测试数据集(25%),使用Boruta算法进行特征选择,并使用SMOTE算法处理类别不平衡。使用五倍交叉验证策略评估模型性能,并使用超参数调整进一步调整最佳模型以增强性能,并使用十倍交叉验证策略进行交叉验证。最终模型是使用 100% 的数据和性能最佳的超参数构建的。(e) 散点区间图描绘了使用接收器操作特征曲线下面积 (AUC-ROC)、准确度、F1 分数、精确度、召回率、Cohen s kappa 和 Matthew s 相关系数 (MCC) 评估的指示模型性能)在十倍交叉验证期间。须线代表 ± SE,粗体水平线表示平均值。
扩展数据图 3 示意图详细说明了构建 AgeXtend 的可解释性和毒性模块模型所涉及的步骤。
(一个) 散点区间图描绘了使用接收器操作特征曲线下面积 (AUC-ROC)、准确度、F1 分数、精确度、召回率、Cohen 的 kappa 和 Matthew 的相关系数 (MCC) 评估的指示模型性能)在十倍交叉验证期间。须线代表 ± SE,粗体水平线表示平均值。值得注意的是,基于签名的生物活性描述符是特征。(乙) 堆积条形图描绘了每个与衰老相关的过程(平均值(|SHAP 值|))在预测类别信息中的贡献。(c) 热图描绘了在 Boruta 选择特征后用于构建指示模型的基于签名的生物活性描述符(签名)的比例。源数据
(一个
) 平均晶须图,显示与衰老相关的生物过程的负面影响因素的老年预测概率的平均值(可解释性模块的第 1 类)。(乙) 热图描绘了 AgeXtend 毒性模块对二甲双胍、牛磺酸和 N-乙酰牛磺酸及其在生物活性潜在空间中最近邻居的预测概率。(c) 热图描绘了二甲双胍、牛磺酸和 N-乙酰牛磺酸与可解释性模块的衰老相关过程的训练数据集中已知激活剂或抑制剂的 Tanimoto 相似系数。(d) 示意图描绘了 AgeXtend 目标模块的结果,突出显示了 mTORC1、HSP90、JAK1 和 HDAC6 的顶部相似配体。(e) 该表包含有关针对 Geroprotectors 筛选的化学数据库的信息。值得注意的是,除了数据库身份之外,还提供了有关数据库版本、预处理前后化合物数量以及高置信度预测的老年保护剂(严格概率截止> = 0.904)的信息。扩展数据图 5 AgeXtend 绘制了潜在的老年保护剂相关化学空间。(一个
) t-SNE 图描绘了在可解释性模块的衰老相关过程的预测概率的潜在空间中,AgeXtend 预测的高置信度老年保护剂(严格概率截止值 >= 0.904)。
数据库中的星号 (*) 表明 t-SNE 代表整个数据库的子集。扩展数据图 6 可解释性模块揭示了 AgeXtend 预测的老年保护剂的抗衰老机制。(一个
) 箱线图描绘了 AgeXtend 上九个突出的衰老相关生物过程的概率分布 (>0.5),表明来自 Aging Atlas 的差异(去)富集代谢物中有老年保护剂。
(乙) 相关散点图描绘了 ELDERMET 肠道微生物群中使用 AgeXtend 的可解释性模块获得的 β 多样性与表观遗传改变 (EA) 概率之间的正相关关系。p 值 (0.049) 使用 Pearson 相关性检验计算得出。(c) 热图描绘了所示内源代谢物的不同化学官能团的相对(去)富集(从上到下) - L-抗坏血酸 (AAA)、脱氧胆酸 (DCA)、3-磷酸-D-甘油酸酯 (DGA),D-葡萄糖6-磷酸(DGP),二羟基丙酮磷酸(DHAP),2-脱氧-D-核糖5-磷酸(DORP),D-核酮糖 5-磷酸 (DRLP)、D-核糖 5-磷酸 (DRP)、D-赤藓糖 4-磷酸 (ERP)、D-果糖 1,6-二磷酸 (FBP)、岩藻甾醇 (FST)、L-组氨酸(HIS)、2-酮戊二酸(KGA)、乙酸盐(ACT)、烟酰胺(NMA)、草酰乙酸(OAA)、吡哆醛 (PAL)、6-磷酸-D-葡萄糖酸酯 (PGA)、吡哆胺 (PMN)、腐胺 (PUT)、核黄素 (RBF)、琥珀酰辅酶 A (SCoA)、三甲胺 N-氧化物 (TNO) 和酵母甾醇 (ZST))。(d) 热图描绘了指示内源代谢物的可解释性模块中所有与衰老相关的生物过程的预测概率。(e) 热图描绘了可解释性模块的衰老相关过程的训练数据集中指定内源代谢物与已知激活剂或抑制剂的 Tanimoto 相似系数。(f) 热图描述了毒性模块对指定内源代谢物的预测概率。(克) 左侧的生存图代表二甲双胍在基于酵母的 CLS 测定中的老年保护功能。误差线代表平均值的标准误差 (SEM)(n≤=≤8 个生物重复)。右侧的箱线图表示使用指定条件的生存曲线下面积 (AUC) 计算的总体生存积分。星号表示统计显着性(p 值≤0.0002,双面 Mann-Whitney U 检验)。对于a和g中的箱线图,中心线显示中位数;框限表示第 25 个和第 75 个百分位数;晶须从第 25 个百分位数和第 75 个百分位数延伸了四分位数范围的 1.5 倍。
扩展数据图 7 在 CLS 测定中,AgeXtend 预测的抗衰老内源代谢物可增强酵母细胞的存活率。
(一个) 箱线图描绘了未处理组 (WT)、载体对照 (Control) 和三种不同浓度的不同生物重复(n≤=≤8 个生物重复)的生存曲线下面积(生存积分)代谢物。中心线显示中位数;框限表示第 25 个和第 75 个百分位数;晶须从第 25 个百分位数和第 75 个百分位数延伸了四分位数范围的 1.5 倍。与各自的车辆对照相比,使用两侧曼-惠特尼 U 检验计算统计显着性。星号表示统计显着性,而 ns 表示无统计显着性。(乙) 生存图描绘了基于酵母的 CLS 测定中 AgeXtend 预测的老年保护代谢物的老年保护作用(n==8 个生物学重复)。误差线代表所有生物重复的存活百分比平均值 (SEM) 的标准误差。底部的箱线图表示使用指定条件的生存曲线下面积计算的生存积分。中心线显示中位数;框限表示第 25 个和第 75 个百分位数;晶须从第 25 个百分位数和第 75 个百分位数延伸了四分位数范围的 1.5 倍。使用两侧曼-惠特尼 U 检验来计算与各自的载体对照相比生存的统计显着性。星号表示统计显着性,而 ns 表示无统计显着性。
扩展数据 图 8 AgeXtend 对老年保护功能具有高命中率预测。
(一个) 生存图描绘了基于酵母的 CLS 测定中 AgeXtend 预测的老年保护代谢物的老年保护作用(n==8 个生物学重复)。误差线代表所有生物重复的存活百分比平均值 (SEM) 的标准误差。底部的箱线图表示使用指定条件的生存曲线下面积计算的生存积分。中心线显示中位数;框限表示第 25 个和第 75 个百分位数;晶须从第 25 个百分位数和第 75 个百分位数延伸了四分位数范围的 1.5 倍。使用两侧曼-惠特尼 U 检验来计算与各自的载体对照相比生存的统计显着性。星号表示统计显着性,而 ns 表示无统计显着性。
扩展数据图 9 在基于酵母的生长测定中,内源代谢物不会引起生长损伤。
(一个) 热图代表使用指定条件下酵母 CLS 测定的生存曲线下面积 (AUC) 计算的生存积分。值得注意的是,在 CLS 测定中使用了三种浓度的 AgeXtend 预测的老年保护代谢物进行治疗。(乙) 用给定浓度的指定内源代谢物处理的酵母细胞(n=-6 生物重复)的生长曲线。误差线代表所有生物重复的归一化 OD600 值的平均值 (SEM) 标准误差。使用两侧曼-惠特尼 U 检验来计算个体生存重复的曲线下面积与相应的载体对照重复相比的统计显着性。星号表示统计显着性(p 值≤<≤0.05),而 ns 表示没有统计显着性。
扩展数据图 10 衰老和非衰老条件下内源代谢物的毒性评估。
(一个) 显微照片描绘了在指定条件下对人成纤维细胞进行苏丹黑 B (SBB) 染色的结果(比例尺 = -100 - µm)。条形图(右)描绘了指定条件下 SBB 阳性细胞数量(n≤=≤10 显微照片)的倍数变化(使用载体对照或未处理进行归一化)。(乙) 该方案(左)描述了用于通过 FDA/PI 测定估计细胞活力的总体实验工作流程。条形图显示了指定条件下的归一化 FDA/PI 值(对于载体对照和测试条件,分别为 n = 4 和 6 个生物重复)。(c) 左侧的方案描述了用于通过 MTT 测定估计细胞活力的总体实验工作流程。条形图显示指定条件下的归一化比色值(对于媒介物对照和测试条件,n≤6 和 9 个生物重复)。(d) 条形图显示了指定条件下的归一化 FDA/PI 值(对于载体对照和测试条件,n = = 10 和 12 个生物重复)。(e) 条形图显示指定条件下的归一化比色值(对于载体对照和测试条件,n = = 11 和 12 个生物重复)。在条形图中(一个—e),误差线代表平均值的标准误差 (SEM),p 值是使用 95% 置信区间的双边 Welch 两样本 t 检验计算的,其中 ns、*、**、*** 和 **** 分别指 p 值 >= 0.05、<0.05、<0.01、<0.001 和 <0.0001。(f) 散点图描绘了基于使用 AgeXtend 计算的预测老年预测分数(x 轴)以及它们与训练数据集中经过实验验证的老年保护剂的最大 Tanimoto 相似性(y 轴)来分离的 DrugBank 化合物的散点图。使用以下方法选择用于实验验证的化合物线虫基于寿命测定的结果突出显示(绿色)。
补充资料
补充表 1-10。
补充表 1 表格表示包含有关用于构建 AgeXtend 的老年预测模块、可解释性模块和毒性模块的化合物数量的详细信息。补充表 2 表格表示包含有关衰老相关生物过程的抑制剂或激活剂的详细信息,用于构建 AgeXtend 的子可解释性模块。补充表 3 表格表示包含有关用于构建 AgeXtend 工作流程的老年预测模块、可解释性模块和毒性模块的化合物的源信息。补充表 4 表格表示,包含有关类别平衡和用于训练老年预测模块、可解释性模块和毒性模块的各种模型的超参数的详细信息。补充表5表5包含来自Signaturers(基于生物活性的描述符)的最终选定特征的签名计数数据,用于构建Geroprediction模块,解释性模块和毒性模块。补充表6表6包含有关用于外部验证的独立数据集的信息。补充表7表7包含有关人肠道微生物组统计数据分析的详细信息。补充表8表8表达包含有关FIM关联,Agextend预测和肠道肠道微生物组队列中的详细信息。补充表9表9包含有关内源性测试代谢物的生物转化器3.0的I阶段和II期生物转化预测的信息。补充表10表10包含有关建立老师预测模型的公开研究的详细信息。值得注意的是,该表还包括有关模型功能,分类器和性能的信息。
权利和权限
Springer Nature 或其许可方(例如协会或其他合作伙伴)根据与作者或其他权利持有者签订的出版协议拥有本文的专有权;作者对本文已接受手稿版本的自行存档仅受此类出版协议条款和适用法律的约束。
关于本文

引用这篇文章
Arora,S.,Mittal,A.,Duari,S。等人。通过可解释的基于人工智能的平台agextend发现Geroprotectors。自然老化(2024)。https://doi.org/10.1038/s43587-024-00763-4
已收到:
公认:
已发表:
DOI:https://doi.org/10.1038/s43587-024-00763-4